Saam Torkan1 ![]() ,
Mohhamad Hossein Sakhaei Shahreza2
,
Mohhamad Hossein Sakhaei Shahreza2
For correspondence:- Saam Torkan Email: Saamtorkan@gmail.com Tel:+98 9133147049
Received: 1 October 2015 Accepted: 16 January 2016 Published: 28 February 2016
Citation: Torkan S, Shahreza MH. VacA, CagA, IceA and OipA genotype status of Helicobacter pylori isolated from biopsy samples from Iranian dogs. Trop J Pharm Res 2016; 15(2):377-384 doi: 10.4314/tjpr.v15i2.22
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the state of vacA, cagA, oipA and iceA genotypes of H. pylori isolated from gastric biopsy samples of dogs.
Methods: A total of 240 gastric biopsy samples were taken from 240 dogs using gastric endoscope. All the samples were cultured and H. pylori-positive samples were analyzed for the presence of various genotypes using polymerase chain reaction (PCR).
Results: Thirteen out of 240 (5.41 %) samples were positive for H. pylori with a higher prevalence of the bacterium in dogs with gastric ulcer (6.36 %). The most commonly detected genotypes were vacA s1a (53.84 %), cagA (46.15 %) and vacA m2 (38.46 %). Total prevalence of iceA1, iceA2 and oipA genotypes were 23.07, 10.8 and 10.81 %, respectively. M2s1a (30.76 %), m1am2 (30.76 %) and m1as1a (23.07 %) had the highest prevalence among other detected combined genotypes. Twenty-two different genotypic combinations were detected with higher prevalence of s1a/cagA+/iceA1/oipA- (23.07 %), m2/cagA+/iceA1/oipA- (23.07 %), and s1b/cagA+/iceA1/oipA- (15.38 %) genotypes.
Conclusion: The presence of similar genotypes in H. pylori strains of gastric biopsy samples of dogs and those of human clinical specimens suggest that dogs may be the source of the bacterium. Proper handwashing and avoidance contact with feces and saliva of dogs are recommended to prevent transmission of H. pylori.
Introduction
Throughout the years, dogs had been the most useful animal companion of humans. Dogs particularly can ease loneliness, reduce allergies, anxiety, depression, stress, and a variety of human diseases, and promote social interaction, encourage exercise and playfulness, and provide unconditional love and affection. In addition, dogs are good hunters, skilled guards, effective narcotic detectors and sharp probes. Company of dogs can increase the level of human immunity against many kinds of zoonotic diseases but there are a lot of diseases that are transmitted from dogs to humans [1,2].
Helicobacter pylori (H. pylori) are gram negative, spiral shaped bacterium, with its main reservoir being humans, particularly the human stomach. The bacterium colonizes most of the population, making it one of the most controversial bacteria in the world that causes peptic ulcer, gastritis, lymphoma, duodenal ulcer, and gastric cancer [3]. Based on the documented data, dogs may play an important role in transmission of H. pylori to humans [4-6]. Gastric Helicobacter infection is common in dogs, with reported high prevalence rates [4-7]. A small percentage (approximately 0.25 – 1.7 %) of human patients suffering from gastric disorders have been diagnosed with gastric Helicobacter spp. that are distinct from H. pylori and morphologically similar to the large spiral-shaped Helicobacter spp. of pet dogs [8]. Contact with dogs has been determined to be a risk factor for human Helicobacter infection [4-8].
To appraise the pathogenicity of H. pylori, evaluation of latent virulence factors and genotypes is essential. The most common but important virulence factors among H. pylori strains of different clinical outcomes of human and animal beings are vacuolating cytotoxin (vacA), induced by contact with the epithelium antigen (iceA), cytotoxin associated gene (cag) and outer inflammatory protein (oip) [9,10]. These genes are usually induced adhesion and invasion to the gastric epithelial cells [11,12]. The vacA belongs to the group of genes with variable genotypes or structures. This gene is associated with injury to epithelial cells. The vacA gene is polymorphic, comprising variable signal regions (type s1 or s2) and mid-regions (type m1 or m2). The s1 type is further subtyped into s1a, s1b and s1c subtypes and the m1 into m1a and m1b subtypes. The mosaic combination of s and m-region allelic types determines the particular cytotoxin and consequently, the pathogenicity of the bacterium [11,13]. The iceA gene has two main allelic variants iceA1 and iceA2 but their functions are not yet clear. Cag pathogenicity island (PAI) has been shown to be involved in persuading inflammation, ulceration and carcinogenesis [12]. The cagA gene has been detected in the specimens taken from severe cases of peptic ulcer [11,12]. The oipA gene plays a significant role in effective colonization of mucosa [11,12]. Genotyping using these virulence markers is considered as one of the best approaches for study of correlations between H. pylori isolates from different samples.
Data on the epidemiology and transmission of H. pylori is extremely significant in order to prevent its distribution and to identify high-risk populations, especially in areas that have high rates of gastritis, peptic ulcers and gastric cancer such as Iran [9,10,13]. Considering the unclear epidemiological aspects of H. pylori in pets and according to the high prevalence of H. pylori all-around the world, the present investigation was carried out in order to study the exact status of vacA, cagA, iceA and oipA genotypes of H. pylori isolated from gastric biopsy samples of healthy dogs and those with gastric ulcer.
Methods
Collection of gastric biopsy samples
From July to September 2014, a total of 240 biopsy samples were collected from the pyloric part of the stomach of dogs of various breeds (male and female, age 7 months to 11 years) that were referred for autopsy at the Department of Small Animal Internal Medicine, College of Veterinary Medicine, Islamic Azad University, Shahrekord, Iran. Samples were collected randomly from healthy dogs (n = 130) and those that had gastric ulcer (n = 110). The dogs with gastric ulcer had a history of gastritis, chronic vomiting and weight loss. Presence of gastric ulcer was diagnosed using the gastric endoscope. Samples were collected in a sterile manner, with care taken to avoid cross-contamination between different sites. Multiple gastric biopsies were taken from each dog. After each sampling, the endoscope and the biopsy forceps were thoroughly cleaned, then sterilized using an activated aldehyde solution. Samples were transported to the laboratory at 4 oC. All samples were kept under refrigeration in plastic bags.
Isolation of Helicobacter pylori
Twenty five milliliters of each homogenized sample were added to 225 mL of Wilkins Chalgren anaerobe broth (Oxoid, UK) supplemented with 5 % of horse serum (Sigma, St. Louis, MO, USA) and colistin methanesulfonate (30 mg/L), cycloheximide (100 mg/L), nalidixic acid (30 mg/L), trimethoprim (30 mg/L), and vancomycin (10 mg/L) (Sigma, St. Louis, MO, USA) and incubated for 7 days at 37 °C with shaking under microaerophilic conditions. Then, 0.1 mL of the enrichment selective broth was plated onto Wilkins Chalgren anaerobe agar (Oxoid, UK) supplemented with 5 % of defibrinated horse blood and 30 mg/L colistin methanesulfonate, 100 mg/L cycloheximide, 30 mg/L nalidixic acid, 30 mg/L trimethoprim, and 10 mg/L vancomycin (Sigma, St. Louis, MO, USA) and incubated for 7 days at 37 °C under microaerophilic conditions. For comparison, a reference strain of H. pylori (ATCC 43504) was employed.
DNA extraction and Helicobacter pylori 16S rRNA gene amplification
Suspected colonies were identified as H. pylori based on the PCR technique. Genomic DNA was extracted from colonies with typical characters of H. pylori using a DNA extraction kit for cells and tissues (Roche Applied Science, Germany, 11814770001) according to the manufacturer’s instructions and its density was assessed by optic densitometry. Extracted DNA was amplified for 16S rRNA gene (primers: HP-F: 5'-CTGGAGAGACTAAGCCCTCC-3' and HP-R: 5'-ATTACTGACGCTGATTGTGC-3') [14]. PCR reactions were performed in a final volume of 50 µL containing 5 µL 10 × buffer + MgCl2, 2 mM dNTP, 2 unit Taq DNA polymerase, 100 ng genomic DNA as a template, and 25 picomole of each primer. PCR was performed using a thermal cycler (Eppendorf Co., Germany) under the following conditions: an initial denaturation for 2 min at 94 °C; 30 cycles of 95 °C for 30 s, 60 °C for 30 s, and 72 °C for 30 s and a final extension at 72 °C for 8 min.
Genotyping of vacA, cagA, iceA and oipA genes of Helicobacter pylori
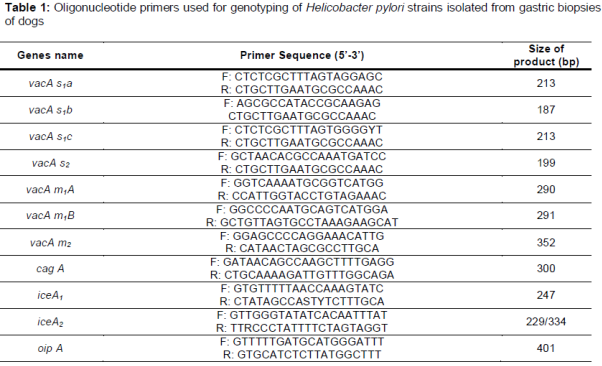
The presence of the oipA, cagA and the genotypes of vacA (s1a, s1b, s1c, m1a, m1b and m2) and iceA (iceA1 and iceA2) alleles were determined using PCR technique. List of primers is shown in [12,15-20].
PCR was performed on 50 μL sample containing 1 μM of each primer, 1 μL of genomic DNA (approximately 200 ng), 1 mM of dNTPs mix (invitrogen), 2 mM of MgCl2, and 0.05 U/μL Taq DNA polymerase (invitrogen). PCR amplifications were performed in an automated thermal cycler (Biometra Co., Germany). The following cycle conditions were used for PCR amplification: for vacA: 32 cycles of 45 s at 95 °C, 50 s at 64 °C, and 70 s at 72 °C; for cagA: 1 min at 94 °C, 1 min at 56 °C, and 1 min at 72 °C; for iceA: 1 min at 94 °C, 1 min at 56 °C, and 1 min at 72 °C and finally, for oipA: 1 min at 94 °C, 1 min at 56 °C and 1 min at 72 °C. All runs included one negative DNA control consisting of PCR grade water and two or more positive controls (26695, J99, SS1, Tx30, 88-23 and 84-183).
Gel electrophoresis and sequencing
PCR amplification products (10 μL each) were subjected to electrophoresis in a 1.5 % agarose gel in 1X TBE buffer at 80 V for 30 min, stained with ethidium bromide, and images were obtained in a UVIdoc gel documentation systems (UK). The PCR products were identified by 100 bp DNA size marker (Fermentas, Germany). In order to confirm the PCR results, the sequencing method was used. For this reason, PCR products of some positive samples were purified with high pure PCR product purification kit (Roche Applied Science, Germany) according to manufacturer's recommendations. Single DNA strands were sequenced with ABI 3730 XL device and Sanger sequencing method (Macrogen, Korea). The result of the sequence of each gene was aligned with the gene sequences recorded in the GenBank database located at NCBI.
Ethical consideration
This work was approved by the ethical committees of Department of Small Animal Internal Medicine, College of Veterinary Medicine, Azad University, Shahrekord, Iran and Research Adjuctancy of the Islamic Azad University of Shahrekord (Ref number 110237IAU). Written informed consent was obtained from the owners of the examined dogs. All stages of sampling was done in such a way as to minimize pain and harm to the dogs.
Statistical analysis
Data were transferred to Microsoft Excel spreadsheet (Microsoft Corp., Redmond, WA, USA) for analysis. Using SPSS 16.0 statistical software (SPSS Inc., Chicago, IL, USA), Chi-square and Fisher’s exact tests were performed, and differences were considered significant at p < 0.05. Distribution of H. pylori genotypes isolated from food stuff was also statistically analyzed.
Results
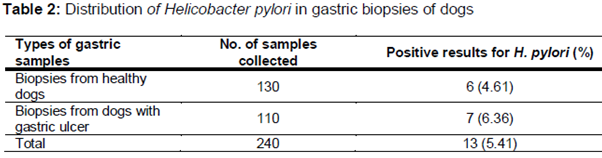
All of the gastric biopsy samples were examined using the culture method. The results of the culture method was confirmed using the PCR technique. shows the total distribution of H. pylori in the gastric biopsy samples of dogs. Of 240 gastric biopsy samples, 13 (5.41 %) were positive for H. pylori. Higher prevalence was seen in dogs with gastric ulcer (6.36 %). There were statistically significant differences amongst the incidence of bacteria and type of samples (p = 0.047).
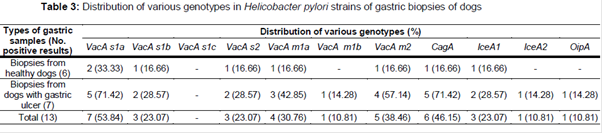
Distribution of vacA, cagA, iceA and oipA genotypes of the H. pylori strains of gastric biopsy samples is shown in . We found that the most commonly detected genotypes were vacA s1a (53.84 %), cagA (46.15 %) and vacA m2 (38.46 %). Total prevalence of iceA1, iceA2 and oipA genotypes were 23.07 %, 10.81 % and 10.81 %, respectively. Significant differences were found between the incidence of s1a and s1b (p = 0.038) and also between iceA1 and iceA2 (p = 0.025), m2 and m1b (p = 0.029), s1a and oipA (p = 0.022) and m2 and iceA2 (p = 0.031).
Distribution of combined genotypes of H. pylori strains isolated from gastric biopsies of dogs is shown in . The most commonly detected combined genotypes were m2s1a (30.76 %), m1am2 (30.76 %) and m1as1a (23.07 %). Significant differences were found between the incidence of m2s1a and m1bs1b (p = 0.020), m2s1a and m1as2 (p = 0.033) and m1am2 and m1as2 (p = 0.031).
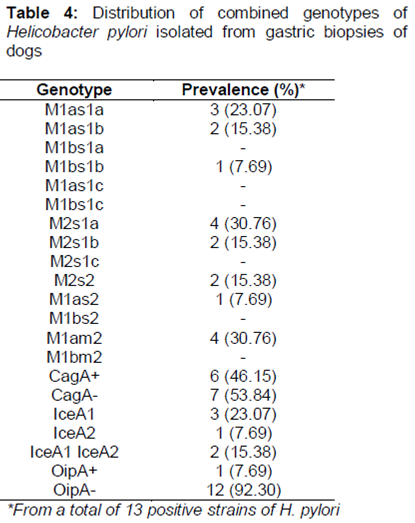
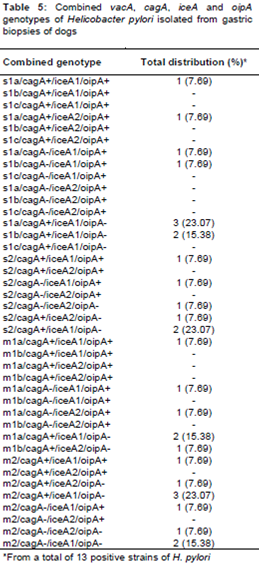
Twenty two different genotypic combinations were detected in the H. pylori strains of gastric biopsy samples (). The most commonly detected combined genotypes were s1a/cagA+/iceA1/oipA- (23.07 %), m2/cagA+/iceA1/oipA- (23.07 %), s1b/cagA+/iceA1/oipA- (15.38 %), s2/cagA+/iceA1/oipA- (15.38 %), m1a/cagA+/iceA1/oipA- (15.38 %) and m2/cagA-/iceA1/oipA- (15.38 %).
Discussion
The human is known as the main host of H. pylori; however pathological examination contrasts with that of many other gastric Helicobacter where the natural host has mild or absent inflammatory response so it seems that H. pylori has originated from another mammalian host in the distant past [21]. The present investigation revealed that dogs, especially those suffering from gastric ulcer, may be the reservoir of H. pylori and/or might be the original host of this bacterium. This finding has been documented previously from all-around the world including Belgium [22], Thailand [23], Italy [4], Denmark [24], Egypt [5] and Iran [6]. The prevalence of H. pylori in dogs from our investigation was 5.41 % which was entirely lower than those of Thailand (17.3 %) [25], Iran (7%) [6] and Italy (78.9 %) [4]. Helicobacter-like organisms are highly prevalent in dogs, with 100 % of random-source dogs [26], 67–100 % of clinically healthy dogs [26-30] and 100 % of laboratory beagles and infected shelter dogs [30,31]. Helicobacter spp. have also been demonstrated in gastric biopsies from 61 – 95 % of dogs [32,33].
The low prevalence rate of H. pylori in our investigation may be due to the fact that the gastric biopsy samples of our study were taken from dogs who were kept in houses. On the other hand, all the dogs in our study had owners. Therefore, they had high levels of health.
As it is obvious the prevalence of H. pylori in household dogs was low. This fact can be proved by two possible reasons: First, household dogs are usually fed healthy and probably cooked foods which were not sources of H. pylori. Second, household dogs are less in contact with stray dogs.
To our best knowledge, the present study was the first report of the genotyping of H. pylori isolated from the gastric biopsy samples of household dogs. As it showed, the most commonly detected genotypes were vacA s1a (53.84 %), cagA (46.15 %) and vacA m2 (38.46 %). CagA-positive strains of the H. pylori have been considered more virulent than cagA-negative strains [34]. In the current survey, some of H. pylori isolated from dogs regardless to clinical outcomes harbor the cagA gene (p > 0.05), therefore, our finding did not support the role of the cagA gene as a predictive marker for increased virulence feature of H. pylori in healthy dogs. Similar deduction can also be made for vacA s1a genotype. According to our findings for the vacA allelles, the vacA s1am2 and vacA m1am2 positive strains were predominant in gastric biopsy samples of dogs. Based on statistical analyses there was a significant correlation between s1a/m2 and m1am2 genotype of H. pylori in healthy and sick dogs. High prevalence of the vacA s1am2 and vacA m1am2 genotypes of H. pylori have been reported previously in the clinical samples of human beings in Iran [35-37]. This may indirectly prove that dogs can be sources of H. pylori for human population. Low prevalence of oipA and iceA showed that these two genotypes were not specific for H. pylori strains recovered from the gastric biopsy samples of Iranian dogs.
Limitations of the study
The high costs of research and also difficulty in convincing owners of dogs to allow samples to be taken from their dogs posed caused some limitations in the number of samples used in this study.
Conclusion
Low prevalence of H. pylori in the gastric biopsy samples of healthy and sick dogs in Iran as well as high presence of vacA and cagA genotypes in both groups of dogs have been detected. The findings also show that Iranian household dogs harbor H. pylori in their gastric tissue similar in genotype to the vacA and cagA allelles of isolates recovered from humans suggesting that dogs may be the ancestral host of the bacterium and that it entered the human population over a period of time. Due to the low prevalence of the virulent strains of H. pylori, basic household hygiene measures such as proper handwashing and avoiding contact with feces and saliva of dogs should be adequate to prevent the transmission of H. pylori.
Declarations
Acknowledgement
References
Archives


News Updates